- Arthropoda (Gliederfüßer)
- Mantodea (Gottesanbeterinnen)
-
Insecta

Äußere Morphologie und Anatomie
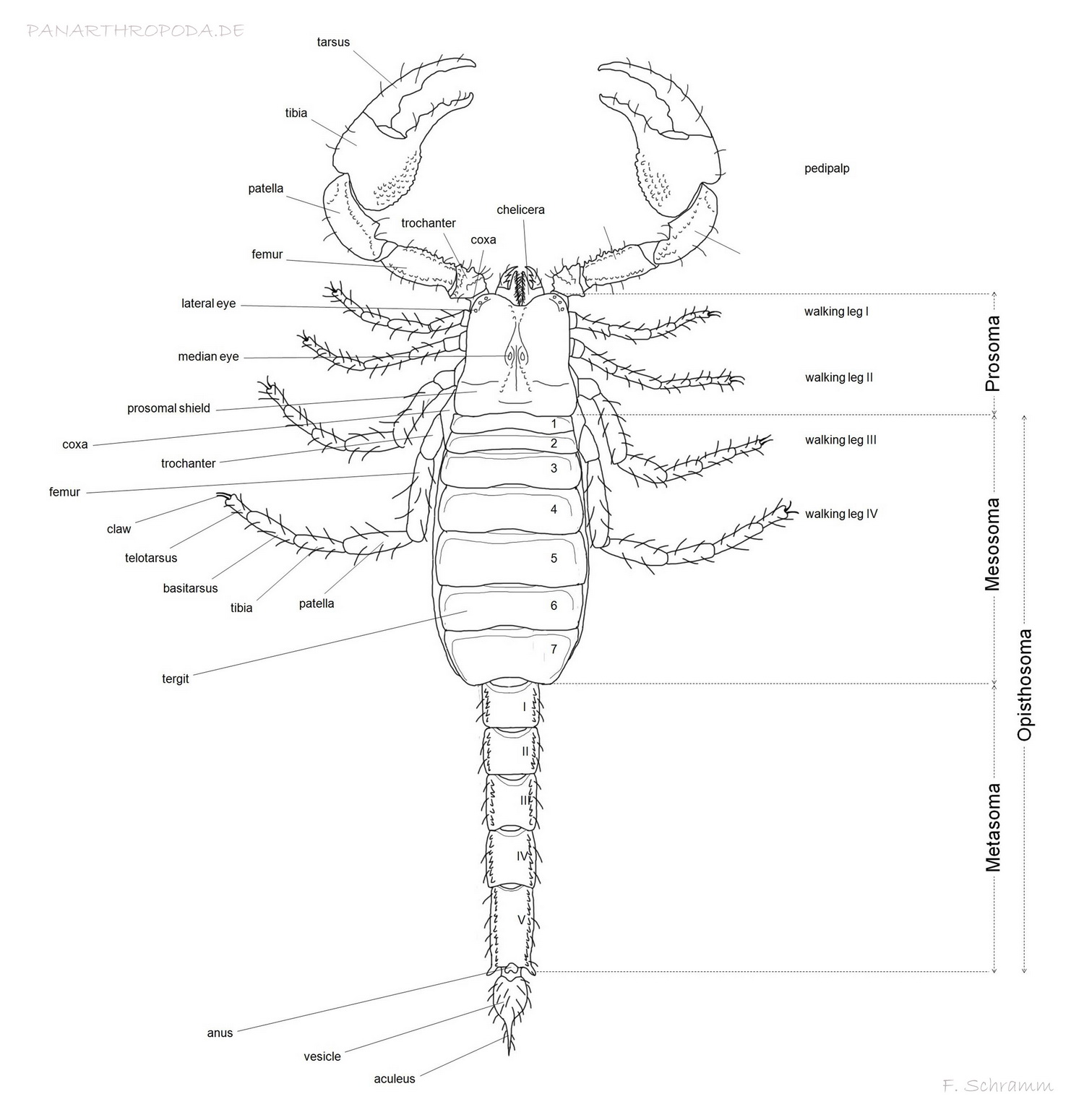
Anatomie des Skorpions im Schaubild
Der Körper der Skorpione ist wie der aller Arachniden in zwei Tagmata unterteilt, das Prosoma und das Opisthosoma (Abdomen). Das Prosoma macht dabei den weitaus kleineren der beiden aus und wird dorsal durch den Carapax bedeckt, welcher (soweit vorhanden) die Median- und Lateralaugen trägt.
Die vorderste am Prosoma beginnende Extremität sind die Chelizeren, dreisegmentige, scherenartige Fresswerkzeuge, die auch anderen Zwecken wie der Reinigung der Extremitäten, indem diese an die Chelizeren geführt werden (Polis, 1990, p11, p12), der Chemorezeption (Abushama, 1964), oder der Lockerung von Substrat beim Graben dienen. Sie befinden sich unmittelbar unterhalb des Carapax, bei helleren Skorpionen ist die Chelizerenmuskulatur mitunter gut durch den Carapax sichtbar. Die beiden sichtbaren distalen Segmente der Chelizeren werden als Tarsus (beweglicher Finger) und Tibia (fester Finger inkl. Basis) bezeichnet. Gewisse morphologische Merkmale der Chelizeren wie u.A. deren Bezahnung fließen mitunter in die taxonomische Eingliederung einiger Skorpiongattungen ein (Vachon, 1963; San Martín, 1972; San Martín & Cekalovic, 1972).
Direkt neben dem sich unter den Chelizeren befindlichen Schlund setzt mit den Pedipalpen (Scherenhänden) das zweitvorderste Extremitätenpaar an. Die Pedipalpen dienen primär des Greifens und der Manipulation von Beutetieren, der Verteidigung und der sensorischen Wahrnehmung (Polis, 1990, p12), spielen aber auch in der Paarung eine sehr wichtige Rolle. Sie bestehen distal beginnend aus Tarsus und Tibia (Beweglicher Finger, fester Finger mit Basis, genannt Manus), welche zusammen die Chela (Scherenhand) bilden, Patella, Femur, Trochanter und Coxa. Die Pedipalpensegmente sind in Abhängigkeit von Familie und Gattung in unterschiedlichster Weise mit Granulierungsreihen (Keilen), sonstigen Granulierungen und Trichobothrien (Becherhaaren) für die Sensorik zur Warnehmung von feinsten Luftbewegungen versehen (Polis, 1990, p12, p36; Stahnke, 1970; Vyas ,1971). Die Anzahl und das Verteilungsmuster dieser Merkmale sind für die Taxonomie von äußerster Bedeutung (Polis, 1990, p17). Besonders die Chelae weisen außerdem in vielen Gattungen Sexualdimorphismen wie verbreiterte oder gestauchte Mani bei Männchen (z.B. Babycurus jacksoni, Scorpio maurus) oder Einkerbungen zum besseren Greifen der weiblichen Chelae bei der Paarung auf (Polis, 1990, p19; Maury, 1975).
Die letzten vier Extremitätenpaare des Prosomas sind die vier Laufbeinpaare. Sie weisen eine zu den Pedipalpen ähnliche Gliederung auf, diese verläuft distal beginnend mit Tarsus inkl. Klauen, Basitarsus und Tibia, Patella, Femur, Trochanter und Coxa. Zwischen Basitarsus und Tibia wie auch Tarsus und Basitarsus können kleine Sporne vorhanden sein (Polis, 1990, p64) (anhand derer z.B. eine leichte Unterscheidung der ansonsten in der Hobbyhaltung oft verwechselten Arten Babycurus jacksoni und Odonturus dentatus möglich ist). Neben dem offentsichlichen Nutzen der Fortbewegung dienen die Laufbeinpaare auch dem Graben und bei Weibchen dem Auffangen der Jungtiere, in dem eine Art Korb gebildet wird (Polis, 1990, p20). Außerdem sind die Beine mit zahlreichen Sinneshaaren und Schlitz-Sensillae versehen und dienen somit in hohem Maße der Sensorik zur Warnehmung von mechanischen und chemischen Reizen (Polis, 1990, p37), wobei der Skorpion durch Gangunterschiede sowohl Richtung als auch Distanz von Vibrationen zu Orten im Stande ist.
Den größten Teil der posterioren ventralen Fläche des Prosomas nimmt die coxo-sternale Region ein, die durch die Coxae des 3. und 4. Laufbeinpaares und das von diesen lateral und anterior eingeschlosse Sternum gebildet wird. Die Form des Sternums im Vergleich zu ausgestorbenen Vertretern der Skorpione weitaus weniger variabel, aber dennoch ausreichend unterschiedlich um ein taxononisches Merkmal für die Familienklassifikation zu bilden (Familien mit zu den anderen deutlich unterschiedlichen Sterna sind z.B. die Familien Buthidae und Bothriuridae) (Polis, 1990, p13). Die Coxae der Laufbeinepaare 1 und 2 sind anterior durch die Coxapophysen verlängert, die zur Bildung eines präoralen Raumes beisteuern (Polis, 1990, p12, p13).
Den größeren Teil des Habitus nimmt das Opisthosoma ein. Dieses besteht aus dem Mesosoma, welches sieben Segmente bis zur Verengung des Körpers umfasst, und dem Metasoma (dem umgangssprachlichen "Schwanz", welcher jedoch keiner ist, da er z.B. Teile des Verdauungstraktes in sich führt und posterior am 5. Metasomasegment im Anus endet) inkl. Telson, das postanal an das letzte Metasoma-Segment anschließt.
Das Mesosoma ist dorsal von sieben teilweise granulierten Tergiten (Rückenplatten) bedeckt. Ventral wird es an den Segmenten 3 - 7 von den Sterniten bedeckt, von denen das 3. - 6. die Stigmenöffnungen (Spirakel) mit den darunter befindlichen Buchlungen aufweisen (Polis, 1990, p15). Das erste Segment weist ventral die vom Genitaloperkulum bedeckte Geschlechtsöffnung auf. Die Form des Genitaloperkulums kann bei einigen Gattungen einen Geschlechtsdimorphismus aufzeigen, so ist dieses z.B. bei Männchen von Pandinus imperator oval, bei Weibchen hingegen herzförmig. Bei den Männchen mancher Arten können außerdem posterior des Genitaloperkulums kleine Papillae vorhanden sein, die entweder leicht sichtbar, oder auch ganz verdeckt sein können. Weibchen weisen dieses Merkmal nicht auf (Polis, 1990, p15).
Das zweite Segment des Mesosoma ist von einer kleinen Basalplatte bedeckt, welche von einem besonderen und nur bei Skorpionen vorzufindendem Extremitätenpaar – den Pecten (Kammorgan) – flankiert wird (Polis, 1990, p15). Diese bestehen aus drei marginalen, zahlreichen medianen Lamellen und kleineren Fulcra und den posterior befindlichen Kammzähnen (Polis, 1990, p20). Die Pecten dienen mit ihren zahlreichen Sinneshärchen der Lamellen und den mit bloßem Auge nicht sichtbaren Sensilla der Kammzähne (Polis, 1990, p23) abermals der Sensorik und nehmen neben Informationen über die Beschaffenheit des Bodengrundes und Vibrationen (Foelix & Müller-Vorholt, 1983) auch chemische Informationen auf. Die Anzahl der vorhandenen Kammzähne variieren zwar je nach Individuum auch innerartlich (Polis, 1990, p20), sie kann aber bei einigen Arten zur Geschlechtsbestimmung herangezogen werden, da zwischen den Geschlechtern teils keine Überschneidungsbereiche der Anzahlen vorhanden sind.
Lateral werden die Tergiten und Sterniten durch die Pleuralmembran zusammengehalten.
Das Metasoma besteht aus fünf Segmenten, die anders als beim Mesosoma nicht von Tergiten und Sterniten bedeckt, sondern rein durch Exoskelettringe umschlossen sind. Auf den Segmenten finden sich wie auch auf den Tergiten Granulierungskeile, die mitunter von taxonomischer Wichtigkeit sind (Polis, 1990, p15). Einige Androctonus ssp. und Parabuthus spp. besitzen auf den dorsalen Flächen von Metasomasegmenten 1 und 2 grobe Strukturierungen, die zur Stridulation in Form von Knackgeräuschen verwendet werden, in dem das Telson über diese gerieben wird (Pocock, 1902, 1904; Millot & Vachon, 1949; Kästner, 1968; Newlands, 1974). Das distale Ende des fünften Segmentes beherrbergt den Anus, direkt daran schließt das zu einem Stachel umgewandelte Telson an.
Das Telson besteht aus dem Versikel (Giftblase) und dem nadelartigen Aculeus. Das Versikel beinhaltet zwei paarig angeordnete Giftdrüsen, die zur Giftabgabe durch Muskeln gegen die Außenwand des Versikels gepresst werden (Polis, 1990, p54).
Quellenangaben
Abushama, F. T. (1964). On the behaviour and sensory physiology of the scorpion Leiurus quinquestriatus (H.&E.). Animal Behaviour 12, 140-153.
Foelix, R. F., and Müller-Vorholt, G. (1983). The fine structure of scorpion sensory organs. II, Pecten sensilla. Bulletin of the British Arachnology Society 6, 68-74.
Kästner, A. (1986). Invertebrate zoology. Vol. 2, Arthropod relatives: Chelicerata, Myriapoda. Tr. H. W. Levi and L. R. Levi, New York: Interscience, 472 pp.
Maury, E. A (1975). Sobre el dimorphismo sexual de la pinza de los pedipalpos en los escorpiones Bothriuridae. Bulletin du Muséum national d'histoire naturelle, Paris, sér. 3, 305: 765-771.
Millot, J. & Vachon, M. (1949). Ordre des scorpions. P.-P. Grasse, ed., Traité de zoologie, Paris, 6, 386-436.
Newlands, G. (1974). The venom-squirting ability of Parabuthus scorpions (Arachnida: Buthidae). South African journal of Medical Science 39, 175-178.
Pocock, R. I. (1902). Arachnida: Scorpiones, Pedipalpi, et Solifugae. Biologia Centrali- Americana (Zoologia), Taylor & Francis, London , 71 pp.
Pocock, R. I. (1902). On a new stridulating organ in scorpions discovered by W. J. Burchell in Brazil in 1828. Annals and Magazine of Natural History, 13, 56-62.
Polis, G.A. (1990). The Biology of Scorpions. Stanford University Press, Standford, California.
San Martín, P. R. (1972). Fijación de los caracteres sistemáticos en los Bothriuridae (Scorpiones). I, Queliceros: Morfología y nomenclatura. Boletín de la Sociedad de biología, Concepción, 44, 47-55.
San Martín, P. R. & Cekalovic, T. K. (1972). Fijación de los caracteres sistemáticos en los Bothriuridae (Scorpiones). II, Queliceros: Estudio de diferenciación a nivel genérico. Boletín de la Sociedad de biología, Concepción, 44, 57-71.
Stahnke, H.L. (1972). Scorpion nomenclature and mensuration. Entomological News 81, 297-316.
Vachon, M. (1963). De l'utilité, en systématique, d'une nomenclature des dents des chélicères chez les scorpions. Bulletin du Muséum national d'histoire naturelle, Paris, 35, 161-166.
Vyas, A. B. (1971). Adaptive diversification of the prosomatic appendages in the scorpion Heterometrus fulvipes. Annals of Zoology 7, 65-80.
F. Schmitz, verfasst 2011