- Arthropoda (Arthropods)
- Mantodea (Mantids)
-
Insecta

Outer morphology and anatomy
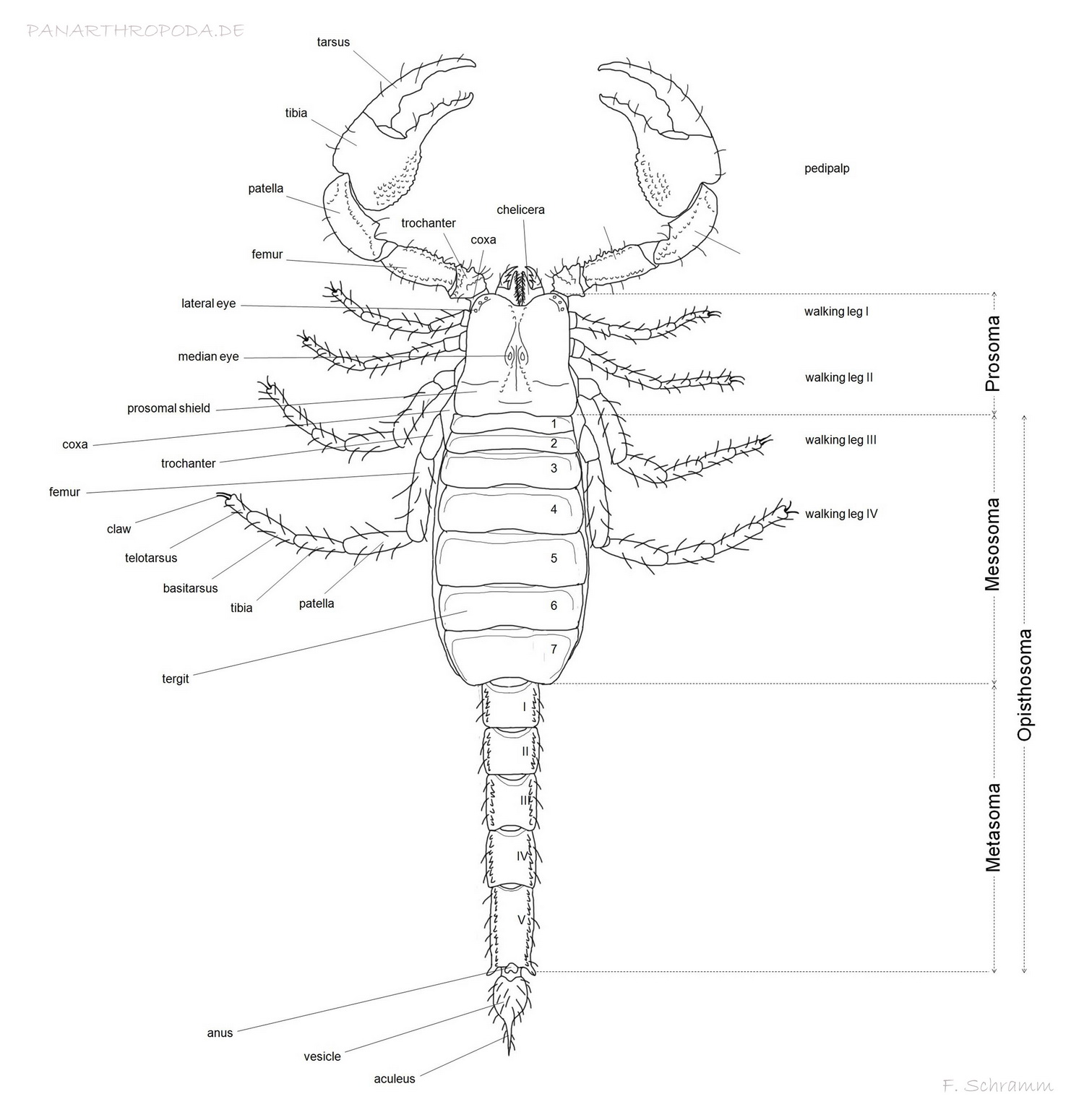
Anatomy of the scorpion
The body of scorpions - like that of all arachnids - is divided into two tagmata, the prosoma and the opisthosoma (Abdomen). The prosoma accounts for the much smaller part of the body and is dorsally covered by the carapace, which (if present) bears the median and lateral eyes.
The foremost extremeties of the prosoma are the chelicerae, three segmented and scissor-like feeding appendages, that are also used for grooming and cleaning other extremeties (Polis, 1990, p11, p12), chemoreception (Abushama, 1964) and even loosening substrat while burrowing. They are located directly underneath the carapace, one can sometimes even see the muscles of the chelicerae shimmer through in ligher colored scorpions. The two visible, distal segments of the chelicerae are the tarsus (moveable finger) and tibia (fixed finger including the base). Certain morphological attributes of the chelicerae are used for taxonomical consideration on the genus level (Vachon, 1963; San Martín, 1972; San Martín & Cekalovic, 1972).
Directly besides the oral cavity (located beneath the chelicerae) the second foremost pair of extremeties, the pedipals, are attached. They primarely serve for grasping and manipulation of prey, defensive purposes and as a sensory organ (Polis, 1990, p12), but are also playing a vital role in mating. Beginning distally they consist of the tarsus and tibia (Moveable finger, fixed finger with base, which is called the manus), which together compose the chela, patella, femur, trochanter and coxa. The pedipalp's segments can depending on family and genus show rows of granules (keels), other granules and different patterns of trichobothria (sensory hairs arising from a cup-like areola), the last of which are used for sensing slightest movements of air (Polis, 1990, p12, p36; Stahnke, 1970; Vyas, 1971). The number and patterns of those characteristics are of great taxonomical importance (Polis, 1990, p17). Especially the chelae exhibit a sexual dimorphism in many species, including broader or compressed mani for males (e.g. Babycurus jacksoni, Scorpio maurus) or indentations in the chelae to assure a better grip on the female's chelae while mating (Polis, 1990, p19; Maury, 1975).
The last four pairs of extremeties located on the prosoma are the walking legs. They exhibit a similar segmentation to that of the pedipalps, distally beginning as tarsus incl. claws, basitarsus and tibia, patella, femur, trochanter und coxa. Between the basitarsus and tibia as well as tarsus and basitarsus little spurs can be present (Polis, 1990, p64) (with the help of which e.g. the in the hobby often confused species Babycurus jacksoni and Odonturus dentatus can be easily distinguished). Apart from the apparent use of locomation the walking legs are also used for digging and in females catching the young during birth by forming a basket (Polis, 1990, p20). Furthermore, the walking legs are heavily endowed with sensory hairs and slit sensilla and are therefore important means of sensing mechanical and chemical stimuli (Polis, 1990, p37), where the scorpion is even capable of sensing distance and direction of vibrations through path differences.
The largest part of the posterior ventral space of the prosoma is taken up by the coxo-sternal region, which is made up of the coxae of walking leg pairs 3 and 4 and the sternum, which on its lateral and anterior borders is enclosed by the before mentioned coxae. The shape of the sternum in comparison to those of former scorpions is not that variable, but nevertheless sufficiently so to serve as a taxonomical characteristic for family classification (Families with significantly different sterna are for example the families Buthidae and Bothriuridae) (Polis, 1990, p13). The coxae of the walking leg pairs 1 and 2 are extended to build the coxapophysis, which serve in closing the preoral cavity ventrally (Polis, 1990, p12, p13).
The greater part of the habitus is taken up by the opisthosoma. It consists of the seven segmented mesosoma and the five segmented metasoma (the colloquial "tail", which is none because it comprises parts of the digestive system which ends posteriorly with the anus on the 5. metasomal segment) including the telson, which is located postanally on the last segment of the metasoma.
The dorsal side of the mesosoa is covered by the mostly granulated tergites (back plates). Ventrally segments 3 to 7 are covored by the sternites, of which the 3rd through 6th bear the spiracles with underlying book lungs (Polis, 1990, p15). The first segment comprises the genital opening, covered by the genital operculum. The shape of the genital operculum can show a sexual dimorphism, e.g. it is of heart-like shape in females of Pandinus imperator and oval in males. Males of many species also exhibit genital papillae on the posterior side of the genital operculum, which are either visible or covered by the latter. In any case, females never exhibit this characteristic (Polis, 1990, p15).
The second segment of the mesosoma is covered by the small basal plate, which is posterially flanked by a very special sensory organ unique to scorpions - the pectines (Polis, 1990, p15). They consist of three marginal and several median lamellae, possibly smaller fulcra and the posteriorly located pectinal teeth (Polis, 1990, p20). The pectines with their almost countless sensory hairs and tiny, invisible to the naked eye, sensillae on the teeth (Polis, 1990, p23) are a major sensory organ for gathering information about the nature of the substrat, vibrations (Foelix & Müller-Vorholt, 1983) and even chemical information. The number of the pectinal teeth present can range widely even among individuals of the same species (Polis, 1990, p20), but can nevertheless be used for sexing individuals in some species, too, when no crossovers of pectinal teeth counts can occur.
Tergites and sternites of the mesosoma are held together laterally by the pleural membrane.
The metasoma consists of five segments, that are in contrast to the mesosoma not covered by tergites and sternites, but by closed exosceletal rings. The segments can show different keels of granules and are therefore of taxonomical importance (Polis, 1990, p15). Some Androctonus ssp. and Parabuthus spp. have severely granulated dorsal surfaces on metasomal segments 1 and 2 which serve as stridulatory organs, with the help of which scraping or buzzing sounds can be created by rubbing the telson over it (Pocock, 1902, 1904; Millot & Vachon, 1949; Kästner, 1968; Newlands, 1974). The distal end of the fifth mesosomal segment comprises the anus, followed by the telson, that has been converted to a stinger.
The telson consists of the versicle and the needle-like aculeus. The versicle containes two paired venom glands, that can be pressed against the inner border of the versicle using muscles for discharging the venom (Polis, 1990, p54).
References
Abushama, F. T. (1964). On the behaviour and sensory physiology of the scorpion Leiurus quinquestriatus (H.&E.). Animal Behaviour 12, 140-153.
Foelix, R. F., and Müller-Vorholt, G. (1983). The fine structure of scorpion sensory organs. II, Pecten sensilla. Bulletin of the British Arachnology Society 6, 68-74.
Kästner, A. (1986). Invertebrate zoology. Vol. 2, Arthropod relatives: Chelicerata, Myriapoda. Tr. H. W. Levi and L. R. Levi, New York: Interscience, 472 pp.
Maury, E. A (1975). Sobre el dimorphismo sexual de la pinza de los pedipalpos en los escorpiones Bothriuridae. Bulletin du Muséum national d'histoire naturelle, Paris, sér. 3, 305: 765-771.
Millot, J. & Vachon, M. (1949). Ordre des scorpions. P.-P. Grasse, ed., Traité de zoologie, Paris, 6, 386-436.
Newlands, G. (1974). The venom-squirting ability of Parabuthus scorpions (Arachnida: Buthidae). South African journal of Medical Science 39, 175-178.
Pocock, R. I. (1902). Arachnida: Scorpiones, Pedipalpi, et Solifugae. Biologia Centrali- Americana (Zoologia), Taylor & Francis, London , 71 pp.
Pocock, R. I. (1902). On a new stridulating organ in scorpions discovered by W. J. Burchell in Brazil in 1828. Annals and Magazine of Natural History, 13, 56-62.
Polis, G.A. (1990). The Biology of Scorpions. Stanford University Press, Standford, California.
San Martín, P. R. (1972). Fijación de los caracteres sistemáticos en los Bothriuridae (Scorpiones). I, Queliceros: Morfología y nomenclatura. Boletín de la Sociedad de biología, Concepción, 44, 47-55.
San Martín, P. R. & Cekalovic, T. K. (1972). Fijación de los caracteres sistemáticos en los Bothriuridae (Scorpiones). II, Queliceros: Estudio de diferenciación a nivel genérico. Boletín de la Sociedad de biología, Concepción, 44, 57-71.
Stahnke, H.L. (1972). Scorpion nomenclature and mensuration. Entomological News 81, 297-316.
Vachon, M. (1963). De l'utilité, en systématique, d'une nomenclature des dents des chélicères chez les scorpions. Bulletin du Muséum national d'histoire naturelle, Paris, 35, 161-166.
Vyas, A. B. (1971). Adaptive diversification of the prosomatic appendages in the scorpion Heterometrus fulvipes. Annals of Zoology 7, 65-80.
F. Schmitz, authored 2011